Decision Making
Animal Communication | Decision Making
Animal Communication
Click on the images to hear the vocalizations.



Our work in the field of animal communication began with the demonstration that chickadee vocalization rates are mass-dependent. This line of research was started with two undergrads in my lab (C. Jackson and A. Schraeder) and was a fairly obvious direction to go from our work on chickadee energy regulation. We looked at the rate at which chickadees sang their “feebeefeebay” song (the top spectragram) and the rate at which they used all other vocalizations (e.g. the “chick-a-dee” vocalization which is illustrated in the middle spectragram and the ‘gargle’ vocalization illustrated in the bottom spectragram). The results showed that song rates increase with an increase in mass, but all other ("non-song") vocalizations actually decreased with an increase in mass. The implication is that non-mating communication is most valuable for "hungry" birds whereas mating communication is most valuable for well-fed birds - an intriguing result. We have since expanded the scope of these studies in two directions. One is an analysis of the structure of the signature "chick-a-dee" call, an interesting vocalization that potentially conveys an extremely rich amount of information to the receiver of the call. The “chick-a-dee” call has a simple syntax, in the sense that it is composed of 4 different types of notes. The birds can include any number of each note type in a call, and can even drop notes from a call. The example illustrated in the spectragram has 2 “A” notes, 1 “B” note, no “C” notes and 6 “D” notes. Given the syntactical properties of the call, chickadees can potentially give an unlimited variety of different chick-a-dee calls, and could therefore potentially encode a fairly large amount of information in this call. In collaboration with a postdoc in my lab (Todd Freeberg - now at Univ. Tennessee), we were able to demonstrate that an unusual variant of the call may be used as a vocalization related to the presence of food (Freeberg & Lucas, 2002). Todd and I published several papers with an undergraduate honors student in my lab (Barbara Clucas -- now at UC Davis) on syntactical aspects of the chick-a-dee call, and on the effect of social inputs on learned components of the call. Much of this work is reviewed in an upcoming book chapter (Lucas & Freeberg, in press).
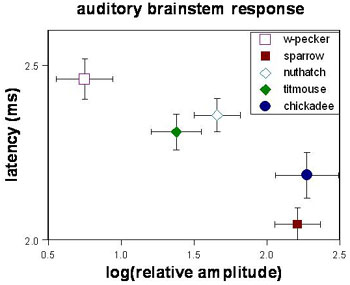
The second direction we are exploring is the result of a collaboration between Todd, me, and two Purdue audiologists (Ravi Krishnan and Glenis Long). This study is a comparative analysis of the auditory acuity of 6 species of birds, and was started when Todd received a training-grant postdoctoral position in the Audiology and Speech Sciences department a few years ago. Our results so far indicate that certain aspects of the activity of the auditory brainstem correlate with the complexity of the vocal repertoire of these birds, whereas other aspects are more closely correlated with taxonomic affinities (Lucas et al. 2002;, Lucas et al. 2007). Moreover, we found seasonal variation in these relationships. This is the first demonstration of seasonal variation in the peripheral auditory system of birds and could potentially be quite important because it expands what we know about the seasonal dynamics of the song nuclei in the avian brain. We also have new evidence that seasonal patterns vary between species: nuthatches show a winter increase in auditory acuity only at 2 kHz, whereas chickadees show a spring increase across a range of frequencies (Lucas et al. in review). Titmice show a spring increase in auditory acuity, but only in a component of hearing related to auditory sensitivity. They do not show seasonality in a component of hearing related to the processing of complex sounds.
I think that this line of research is important because we are starting to address aspects of avian vocal communication from both the perspective of the sender of auditory signals and also from the receiver's perspective. We have developed the brainstem recording technique enough that we can start to tie together what we know about the chickadee vocal repertoire with an analysis of the neurophysiology of the chickadee peripheral auditory system. Quite unexpectedly, our neurophysiological work has also raised some questions about seasonal variation in auditory acuity that suggest that a closer analysis of seasonal variation in signal production would be worth our efforts.
Decision-Making
Energy regulation

One of the most important recent advances in the study of the evolution of behavior has been a reevaluation of the role of physiological state (such as the level of fat reserves) in decision-making processes. The addition of state to the study of behavior greatly increases the complexity of the problem of trying tunderstand the evolutionary context of behavior. We model behavior by modeling the costs and benefits of the behavioral alternatives available to an animal, assuming that the animal will chose the option with the highest net benefit. These models become much more complicated when state dependence is added, because any decision an animal makes will have delayed effects on the physiological state of that animal and therefore on the suites of behaviors affected by physiological state. Under many circumstances, these delayed future effects cannot be modeled analytically. Unfortunately, we cannot ignore these issues: physiological state clearly has a major impact on behavior. For example, our work shows that hungry birds are more impulsive in their diet-choice decisions than are sated birds, that food-storage decisions are strongly affected by a bird's body mass, and that both social and environmental conditions can in turn affect the regulation of body mass. Clearly there is a fairly complex feedback between environment, behavior, and physiological state.
A major breakthrough in the study of state-dependent behavior (or behavior that changes with physiological state) has been the development and application of dynamic optimization techniques, and specifically dynamic programming. While the theory is actually fairly old (it was developed by R. Bellman in the 1950's), only recently has it been used extensively in the study of behavior. My time-constraint models of diet choice provided a fairly early example of the application of this technique. Since then, we have focused on food hoarding behavior and have developed a comprehensive theoretical treatment of energy regulation in small birds.
A number of species will cache food for future use instead of eating it immediately on capture. Previous research has shown that an important function of caching behavior is that it reduces variation in resource availability in highly variable environments. Environmental variability, in turn, is particularly important for small animals, because the amount of time an animal can persist on fat stores is size-dependent. For example, chickadees weigh only 10 g and cannot survive 24 h in the winter on their fat stores. This makes cached food the only long-term form of energy storage available to these birds. Tight energy budgets in these birds also make the fitness consequences of caching behavior fairly transparent, which in turn makes this an excellent model system.
An animal signals the time horizon of its decision (i.e., how far into the future the animal considers the consequences of its decisions) through its choice of how long it leaves food in a cache site before retrieving it. This provides a unique insight into the temporal dimension of this foraging decision; few other behaviors have this trait. We have shown that caching decisions are indeed strongly affected by physiological state. In titmice, the state-dependent properties of caching behavior can differ qualitatively from those of diet choice: compared to thin birds, fat animals are impulsive in their caching decisions (i.e., they do not cache) but are not impulsive in their diet choice decisions (i.e., they are more likely to ignore low quality prey in order to wait for higher quality prey). These results suggest that animals treat future payoffs differently for these two types of decisions.
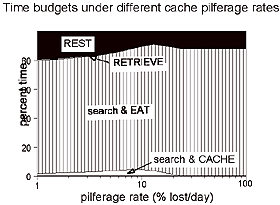
In chickadees, we showed that state-dependent changes in caching decisions covary with state-dependent changes in the overall time budget. The implication is that changes in caching behavior are a part of a suite of behavioral changes that occur when energy levels vary. This underscores the need for broad scale models of behavior that incorporate realistic interactions between different types of behavior. We have taken this approach in developing models of caching behavior. Our theoretical results suggest that the value of foraging-related rewards should be both state-dependent and dependent on environmental quality. In high-quality environments, birds should weigh foraging-related requirements more heavily than non-foraging rewards when they are thin. In low-quality environments, foraging-related requirements should be given preference to non-foraging requirements at all weights; however, the time horizon of foraging decisions should be shorter if the animal is light weight (i.e., it should accept any immediate reward). The net result that the state-dependent properties of caching behavior should depend on the type of environment a bird occupies. Our results support this prediction. The relationship between caching behavior and body size changes with overall environmental quality: in a poor-quality environment, chickadees cache less when light weight, but in a high-quality environment they cache more when light weight.
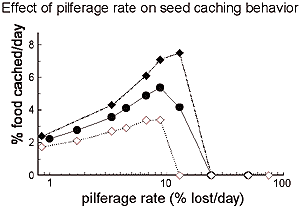
If the cache is pilfered, we showed that birds compensate for the loss of the stored food by caching more food, a result contrary to a number of published predictions including our own. We re-evaluated this prediction with a new dynamic program and showed that the prediction is incomplete. Instead, we showed that caching rates should be a unimodal function of pilferage rate: increased pilferage rates when absolute pilferage is low should result in higher caching rates (because the cache has an intrinsic value that can be compensated for with higher caching rates), but increased pilferage rates when absolute pilferage is high should result in lower caching rates (because the marginal cost of adding seeds to the cache is too high when those seeds are very likely to be stolen before they are retrieved). I think that this is an excellent example of how complex behavioral decisions can result from the three-way feedback described above that is embodied in dynamic decision-making systems. It also underscores the utility of the modeling/empiricism cycle.
Our empirical results have turned up intriguing seasonal aspects to energy regulation. There appears to be a seasonal component to the expression of caching behavior, even in the laboratory under constant conditions. In the winter, chickadees cache more food and eat more seeds (as opposed to insect food) than in the summer. In response to a prolonged shortage of food, birds in winter adjust caching rates so that they can maintain fairly stable weights, but birds tested in the summer do not do this. In addition, an experimental analysis of the effect of seed pilferage on energy regulation showed that birds compensate for seeds stolen in the winter by caching more seeds, but that this compensation does not occur in the summer. The fact that energy regulation patterns differ across seasons suggests that there are fitness consequences to the typical winter regulation patterns that are not adaptive in the summer. It is unclear exactly what these consequences are because there are a host of factors that vary seasonally, from simple time constraints (due to day length) to varying reproductive requirements. With funding from NIMH, we just started a study of seasonal aspects of energy regulation in chickadees and how these seasonal energy cycles correlated with hormone profiles (corticosterone and testosterone), spatial memory and hippocampal structure and function. This is an exciting next step for my lab because it provides insight into the mechanisms of energy regulation in a way that is a perfect complement to the adaptive aspects of energy regulation that we have been studying for some time.
Dynamic game theory
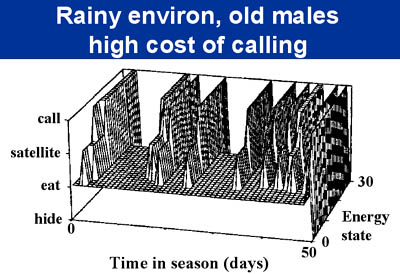
One exciting direction that we are pursuing in our modeling efforts is the development of stochastic dynamic games, where both physiological-state dependence and frequency- and density-dependence can be incorporated into the model. My feeling is that both phenomena are nearly ubiquitous, although dynamic game theory has been underdeveloped to date. I collaborated with Dr. Richard Howard (his graduate student) on some dynamic games of calling behavior in anurans. Male anurans show at least two alternative mating tactics: some will advertise to females by calling, while others (satellites) sit silently near the calling males and intercept females attracted by the callers. A fair amount of work has been published on the relative mating success of both tactics, so we have a good empirical base for the model. These papers were the first that I know of to show how both frequency-dependence and density-dependence can be incorporated into this class of models. Previous dynamic games typically only evaluated the effects of frequency-dependent payoffs on the expression of behavior. In frogs, for example, the relative frequency of satellites in the population will affect the mating success of callers. However, density-dependence is also important; in fact, density-dependence has been the cornerstone of much of ecological theory (e.g., competition theory and predator-prey dynamics). In anuran mating systems, for example, predation risk is likely to vary with the density of the chorus. The ability of the males to attract females may also depend on chorus size. Given that density-dependence is so common in ecological systems, this is likely to be an important direction for future research
I also collaborated with another colleague, Dr. Peter Waser and his student Scott Creel on a dynamic model of dispersal behavior in dwarf mongooses. We showed that dispersal decisions in these mongooses are affected by fairly subtle delayed effects of movement and dominance status; this is a perfect system for the use of dynamic optimization. We also devised a method of incorporating inclusive fitness into dynamic programs; this is the first description of which I am aware of a model of this type.
In collaboration with Dr. Robert Gibson (U. Nebraska) and Adam Boyko (a graduate student in my lab), we published a dynamic model of lekking behavior with an eye to understanding what role predation risk plays in the development of this intriguing mating system. Finally, in collaboration with Ed Harris (a graduate student now in Scotland), we studied the dynamic aspects of sperm competition in salamanders.