
B. Nitrogen Fixation In a Unicellular Cyanobacterium
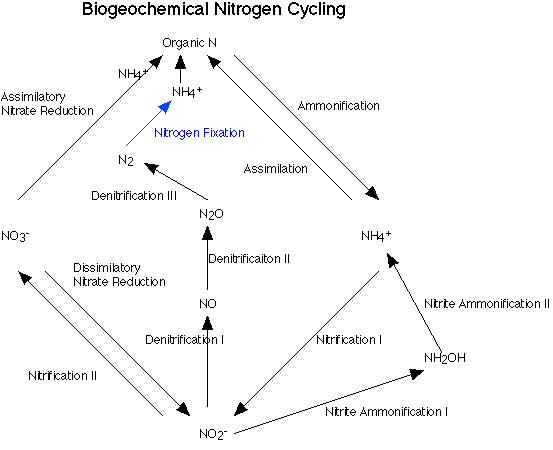
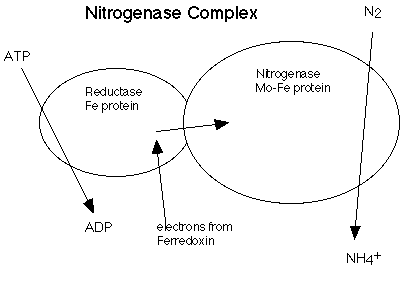
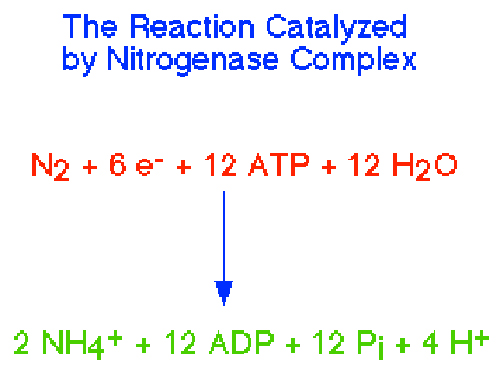
1. Strategies to Protect NitrogenaseThe biological conversion of atmospheric dinitrogen (N2) to ammonia (NH3), a process called dinitrogen fixation, is central to global nitrogen cycles (Fig. 7). An enzyme complex called nitrogenase (Fig. 8) catalyzes this reaction in some cyanobacterial and bacterial species. Nitrogenase performs the fixation reaction , (Fig. 9) a six electron reduction of N2 to NH3, using 6 NADPH and as many as 12 ATP. The nitrogenase complex can be irreversibly inactivated by atmospheric oxygen (O2). The sensitivity of nitrogenase to oxygen may be a remnant of earlier times, billions of years ago, when the Earth's atmosphere was lower in oxygen. Many of the organisms that can fix dinitrogen can only do so in environmental niches where oxygen is either absent or at very low levels.
{kind=link}
{kind=link}
{kind=link}
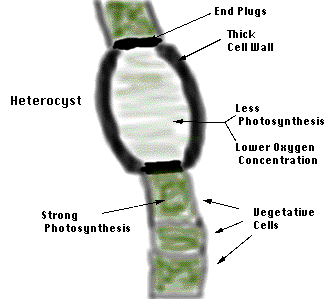
By their nature, cyanobacteria produce oxygen when water is split during photosynthesis. Thus, it is surprising that some photosynthetic cyanobacteria can also fix dinitrogen. The N2-fixing, or diazotrophic (literally dinitrogen-eating), cyanobacteria have developed interesting methods to protect their oxygen-sensitive nitrogenase from both atmospheric and photosynthetic oxygen. Some cyanobacteria that form filaments (strings) of cells develop specialized cells that perform dinitrogen fixation, called heterocysts. The heterocysts form a thickened cell wall (glycocalyx) and plugs at the junction between cells in a filament (Fig. 10) to limit the diffusion of atmospheric oxygen into the heterocyst. Photosynthesis is turned off in heterocysts and they survive on sugars supplied by the other cells of the filament. These other cells, called vegetative cells, are repaid for their sugars with fixed nitrogen compounds made by the heterocyst.
{kind=link}
Other cyanobacteria use different strategies to protect their nitrogenase from oxygen. Certain filamentous strains form bundles of filaments. In these bundles, only certain filaments will fix dinitrogen, the other filaments somehow shielding the N2-fixing filaments from oxygen. Single celled cyanobacteria that do not form colonies cannot use the same strategies as the filamentous and colony-forming organisms. The unicellular cyanobacteria seperate in time the functions of photosynthesis and dinitrogen fixation. In the natural environment, these organisms photosynthesize and produce oxygen in the light during the day, and then fix dinitrogen in the dark at night.
Dr. Sherman's laboratory has been studying the temporal separation of photosynthesis and dinitrogen fixation in the unicellular diazotrophic cyanobacterium, Cyanothece sp. strain ATCC 51142.
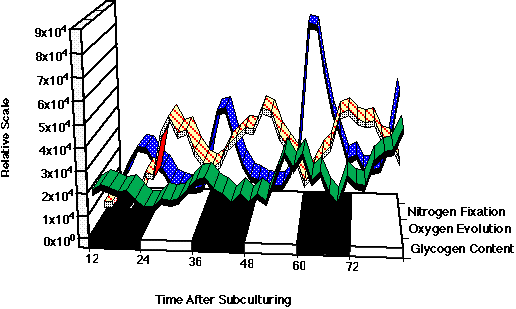
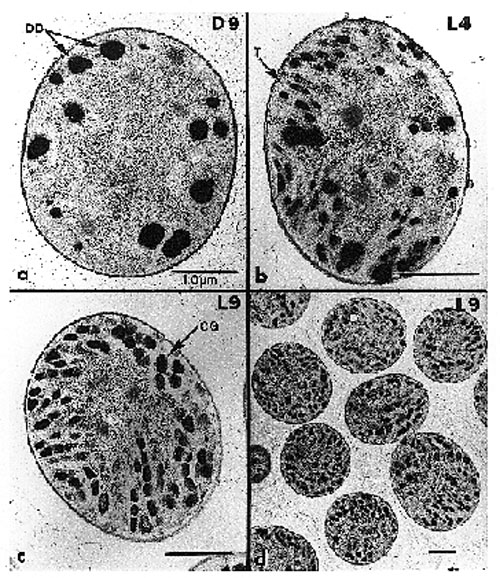
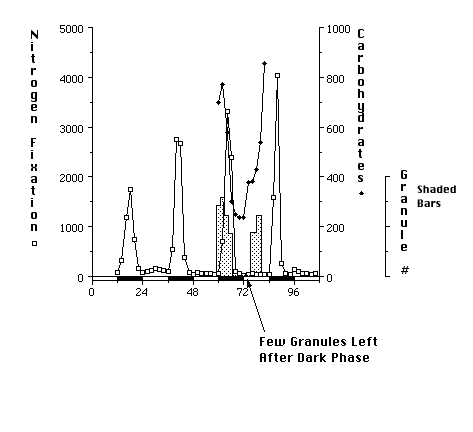
2. Metabolic Rhythms and Regulation in CyanotheceCyanothece sp. strain ATCC 51142 is a marine, unicellular, diazotrophic cyanobacterium. It appears that the strategy of temporal separation is used to protect oxygen-sensitive nitrogenase from photosynthetic oyxgen. When grown under alternating periods of 12-h light and 12-h dark (LD), photosynthetic oxygen evolution is limited to the light phase (Fig. 11). The fixation of dinitrogen occurs during discrete periods in the dark phase. Dinitrogen fixation is a rather energy-expensive process, however, in the dark, photosynthesis cannot directly supply the energy and reducing power needed. In Cyanothece sp., it appears that dinitrogen fixation in the dark is powered by stored carbohydrates. During the latter part of the light phase, carbohydrates accumulate in large granules that form between the photosynthetic membranes (thylakoids) (Fig. 12). The carbohydrate granules can be visualized in the electron microscope. The number of granules and the amount of carbohydrate are highest just prior to the onset of the period of dinitrogen fixation (Fig. 13). Most of the carbohydrate is degraded to fuel dinitrogen fixation, so there are very few granules remaining at the end of the dark phase (Fig. 14). The conversion of carbohydrates to energy and reducing power is a process called respiration and requires oxygen. Thus, as the carbohydrates as used, oxygen within the cell is also being used. This in effect lowers the oxygen concentration within the cell and helps to protect nitrogenase from inactivation by oxygen.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
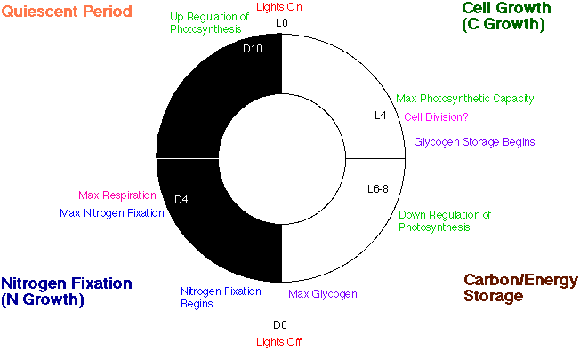
In the natural environment, there are daily periods of light (the day) and dark (the night). In the laboratory, environmentally controlled growth chambers can be used to grow cyanobacteria under different patterns of illumination (photoperiods). Cyanothece sp. can be grown under conditions of continuous illumination. Since there are no longer any dark periods, when will cells grown under continuous light fix dinitrogen? As with the culture under LD conditions, dinitrogen fixation occurs in discrete periods. The timing of these peaks corresponds to the dark period of DL cycles, but since this experiment was conducted with continuous light, this period is called the subjective dark phase. Thus, 16 hours after subculturing, there is a peak of dinitrogen-fixation activity whether the culture is grown under LL or LD conditions. It is interesting to note that even under LL conditions, the capacity for photosynthesis is reduced during the period of dinitrogen fixation. Furthermore, even though light energy is available, the cells still appear to use accumulated carbohydrates to fuel dinitrogen fixation. It seems likely that the carbohydrates are needed so that during dinitrogen fixation there is intense respiration, thus protecting nitrogenase from oxygen. A model of the circadian patterns is shown in Figure 15.
{kind=link}
Growth under conditions of continuous dark (DD) is not a natural activity for cyanobacteria. Many species of cyanobacteria will not grow without light. However, Cyanothece sp. can be grown under DD conditions when provided with a source of food. In this case, the carbon source used was glycerol. Will cultures under DD conditions show metabolic cycles? Will cultures provided with all the food they need accumulate carbohydrates? With a DD culture, questions like these can be answered. Preliminary results indicate that under DD conditions, dinitrogen fixation occurs during discrete periods of the subjective dark phase. Photosynthetic capacity is reduced during the period of dinitrogen fixation. And, carbohydrates are indeed accumulated during the subjective light phase and dissipated during the period of nitrogen fixation. This is very interesting since these cells are supplied with enough glycerol to provide a continuous stream of energy for dinitrogen fixation. Instead of fixing dinitrogen continuously, the cells convert the glycerol into carbohydrates and store these for use in fueling dinitrogen fixation during discrete periods of a daily cycle. As with the LL and LD cultures, it seems likely that the respiration of the stored carbohydrates is needed to lower oxygen concentrations in the cell and protect nitrogenase from inactivation.
How does the cell control these metabolic cycles? Under LD conditions, it is easy to say that the changes from light to dark or vice versa are the signals that control the cycles. Under either LL or DD conditions, there are no such signals. The control of the cycles in Cyanothece sp. may be similar to processes observed in other organisms that have been called circadian rhythms. Jet lag is one manifestation of circadian rhythms in humans. The mechanisms of circadian rhythms are not clear and a molecular clock has yet to found. It is possible that the rhythms in Cyanothece are in part correlated with changes in the redox state of the cell through the action of thioredoxin.
To learn more about this work, follow this link.
This work was performed in part by Dr. Mark A. Schneegurt.
Dr. Mark Schneegurt's Homepage
Back to Sherman Lab Homepage