
J. Microarray Analysis of the Genome Wide Response to Iron Deficiency and Iron Reconstitution. Plant Phys 03
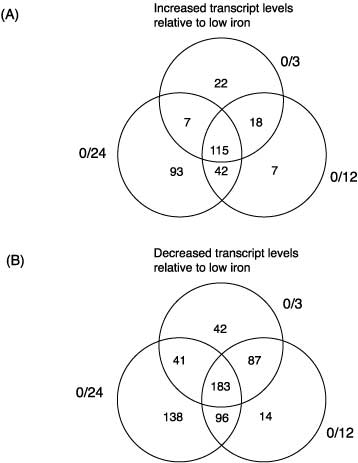
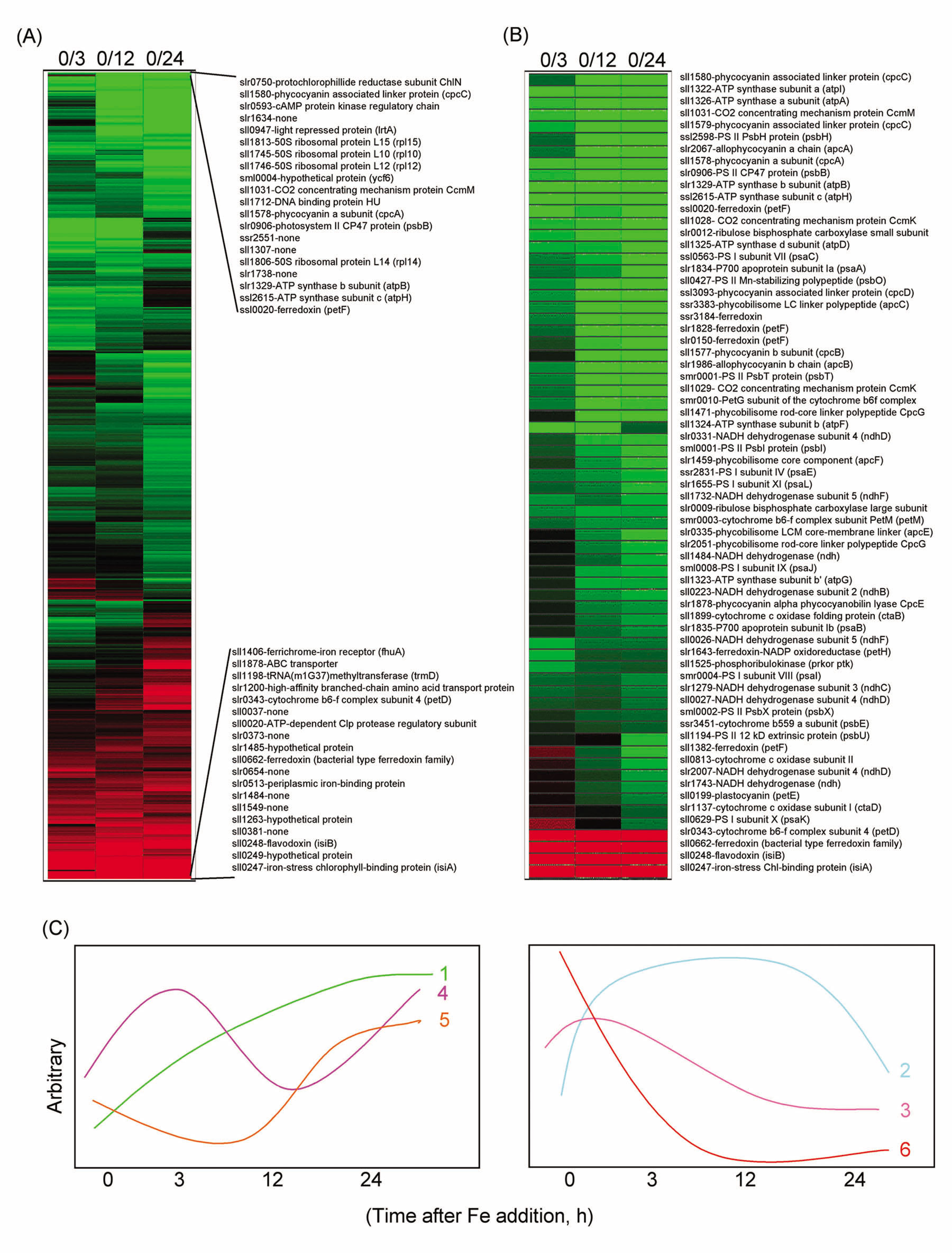
1.) SummaryA full-genome microarray of the (oxy)photosynthetic cyanobacterium Synechocystis sp. PCC 6803 was used to identify genes that were transcriptionally regulated by growth in iron (Fe)-deficient versus Fe-sufficient media. Transcript accumulation for 3,165 genes in the genome was analyzed using an analysis of variance model that accounted for slide and replicate (random) effects and dye (a fixed) effect in testing for differences in the four time periods. We determined that 85 genes showed statistically significant changes in the level of transcription (P _ 0.05/3,165 _ 0.0000158) across the four time points examined, whereas 781 genes were characterized as interesting (P _ 0.05 but greater than 0.0000158; 731 of these had a fold change _1.25_). The genes identified included those known previously to be Fe regulated, such as isiA that encodes a novel chlorophyll-binding protein responsible for the pigment characteristics of low-Fe (LoFe) cells. ATP synthetase and phycobilisome genes were down-regulated in LoFe, and there were interesting changes in the transcription of genes involved in chlorophyll biosynthesis, in photosystem I and II assembly, and in energy metabolism. Hierarchical clustering demonstrated that photosynthesis genes, as a class, were repressed in LoFe and induced upon the re-addition of Fe. Specific regulatory genes were transcriptionally active in LoFe, including two genes that show homology to plant phytochromes (cph1 and cph2). These observations established the existence of a complex network of regulatory interactions and coordination in response to Fe availability.
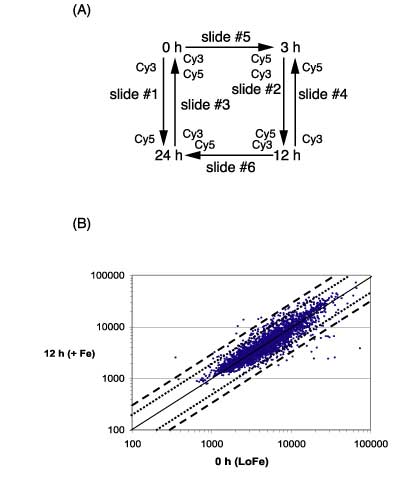
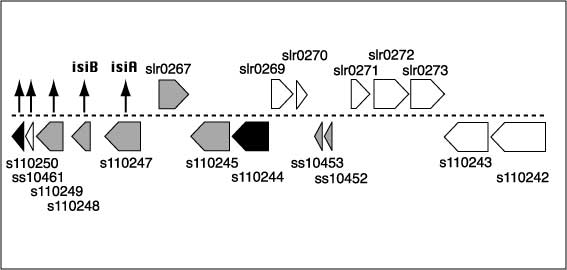
2.) IntroductionFe is an essential element that is required for the growth and development of all organisms, including microorganisms (Hantke, 2001) and plants (Thimm et al., 2001; Negishi et al., 2002). Although Fe is abundant in nature, the availability of this element is very limited because of its poor solubility in aerobic environments. In the presence of oxygen at physiological pH, the rapid oxidation of the ferrous form to the ferric form leads to the precipitation of Fe and its essential unavailability. Thus, living organisms have developed various mechanisms to solubilize Fe to improve its bioavailability (Fox and Guerinot, 1998; Ratledge and Dover, 2000). Fe is of great importance for the growth of both pathogenic and nonpathogenic bacteria, and many strains devote a significant portion of their genome to the regulation of and the acquisition of Fe (Earhart, 1996; Paustian et al., 2001). Cyanobacteria are (oxy)photosynthetic organisms in which Fe stress has been studied in some detail (Straus, 1994; Behrenfeld and Kolber, 1999). Fe deficiency results in a variety of physiological and morphological changes in cyanobacteria, the most obvious of which is a significant change in cellular pigmentation. The overall changes include: loss of the light-harvesting phycobilisomes (Guikema and Sherman, 1983), changes in the fluorescence and absorption characteristics (Guikema and Sherman, 1983, 1984; Pakrasi et al., 1985a, 1985b), reduction in the number of thylakoids (Sherman and Sherman, 1983), and replacement of proteins with cofactors containing Fe to those with non-Fe cofactors, such as ferredoxin with flavodoxin (Laudenbach and Straus, 1988; Laudenbach et al., 1988; Straus, 1994). Most importantly, a novel chlorophyll (Chl)-binding protein, encoded by isiA, is synthesized (Pakrasi et al., 1985b; Laudenbach and Straus, 1988; Burnap et al., 1993). This IsiA protein resembles CP43 (it is sometimes termed CP43) and has recently been shown to form an 18-mer around the PSI trimer (Bibby et al., 2001; Boekema et al., 2001). This gene is also regulated by salt concentrations (Vinnemeier et al., 1998) and may be the ancestor of the Prochlorococcus sp. Chl a2/b2 light-harvesting protein (Ting et al., 2002). Many other changes are associated with Fe deficiency in cyanobacteria and cells continue to grow, although the growth rate is somewhat lower and the cells are smaller. The addition of Fe to the cultures can reverse these changes, and cells return to normal within 18 to 24 h after the addition of Fe (Sherman and Sherman, 1983; Riethman et al., 1988). Microarray technology permits an assay of global gene expression patterns under a variety of experimental conditions. These arrays are particularly efficient in organisms for which the entire genome has been sequenced, such as Synechocystis sp. PCC 6803, which is now thought to have 3,264 genes (Kaneko et al., 1996; see Cyanobase at http://www.kazusa.or.jp/cyano/cyano.html). Microarrays have been developed for many systems, including for bacteria such as Escherichia coli (Richmond et al., 1999; Tao et al., 1999; Arfin et al., 2000) and for plants such as Arabidopsis (Pérez-Amador et al., 2001; Seki et al., 2001). A series of papers have appeared utilizing Synechocystis sp. PCC 6803 microarrays (Hihara et al., 2001; Suzuki et al., 2001; Gill et al., 2002; Kanesaki et al., 2002). These arrays have been used to monitor changes in different environmental parameters. The arrays that we constructed, in conjunction with the laboratory of Dr. Rob Burnap (Oklahoma State University, Stillwater), contain (in triplicate) cDNAs up to 2 kb of the 3,165 genes annotated in the Kazusa sequence before May 2002. The substantial pigmentation changes under Fedeficient growth provide an easy way to determine the cellular response to Fe deficiency or the redevelopment of the normal phenotype. Thus, Fe deficiency is an ideal system in which to study global gene expression in cyanobacteria. In a previous study, we developed a differential expression using customized amplification library for the analysis of global gene expression in the unicellular cyanobacterium, Synechocystis sp. PCC 6803 (Singh and Sherman, 2000). We now extend this study through an analysis of a full-genome microarray of Synechocystis sp. PCC 6803. We identified transcriptional changes in many genes that code for proteins involved in assembly or disassembly processes (e.g. chaperones and proteases) and in the structural proteins (e.g. IsiA or phycobiliproteins). The arrays also enabled us to detect genes involved in the regulation of these processes and for those that encode proteins needed for the acquisition and storage of Fe (Katoh et al., 2000, 2001). In this study, we identify many genes that are transcriptionally regulated during Fe deficiency and after the re-addition of Fe and that provide new insights into optimization of biological processes that enable cells to grow during nutrient limitation.
Links to Figures in the paper:
Figure 1
Figure 2
Figure 3
Table 3
Figure 4
{kind=link}
{kind=link}
{kind=link}
{kind=link}