
C. SYNECHOCYSTIS MUTATIONS AND A STRUCTURAL MODEL OF MSP
1. Random Mutantsa. Digital Imaging Spectrometer (DIS)
Imaging spectroscopy is defined as the combined analysis of both spatial and spectral information so that each picture element (pixel) in a 2-D scene includes a third dimension of spectral information. This means that the absorption spectrum of any pixel or group of pixels within an image can be measured. The KAIROS ColonyImager DIS (Yang and Youvan, 1988; Yang, 1994; Youvan, 1991) utilizes a charged couple device (CCD), detectors and diffused light sources in both a transmission and reflectance mode (Fig. 16). The detector is a 16 bit CCD camera with: 1) a large dynamic range of light sensitivity; 2) linearity in photon conversion to electron signal; 3) geometric stability; and 4) the ability to integrate photons over an extended period of time which yields greater sensitivity in low-light applications. The DIS can be used in either reflectance or transmittance mode, depending on the needs. It can also be used for fluorescence by selecting an appropriate long pass filter and this is the mode in which we use the machine most frequently. The DIS image processing capabilities permit us to generate apparent quantum yield information by digitally dividing a fluorescence image with a gray scale-inverted absorption image.
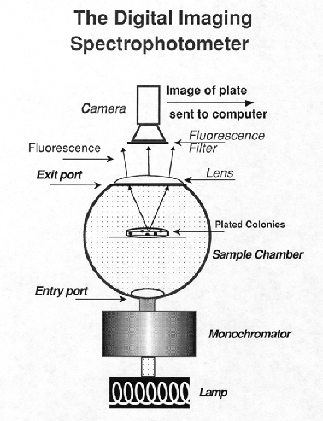{kind=link}
We devoted much effort to modifying and refining the equipment and the procedures. We were aiming to automate the screening, such that a high percentage of colonies selected were indeed mutants. We have delved into minute details, such as the precise colony size on the plates, the physiological state of the colonies (i.e., state transitions), the standard deviation of fluorescence intensities of colonies within a plate, and plate-to-plate variations of the mean intensity, etc. We have finally developed a procedure that is reproducible, minimizes variations, and is relatively rapid (Meunier, 1997b).
b. Random Mutants of psbOThis procedure enabled us to isolate quite a few mutations in MSP. The most surprising aspect of this mutant search was that we obtained single amino acid mutations throughout the protein, from the N-terminus through amino acid 219. Some of the mutations demonstrated subtle changes in photosynthetic properties, such as oxygen evolution or PC/Chl. Some mutations had altered state transitions and others had changes in the amount of F696 at 77 K. Some mutations, such as MSP11 (P219L) acted like the DpsbO mutation we had constructed previously. Thus, we obtained mutations from throughout the protein with the full gamut of phenotypes related to psbO mutations.
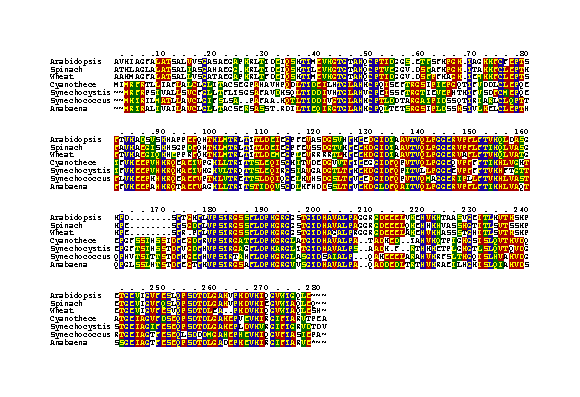
Despite this technical success, the overall analysis of the mutants was quite difficult. From the time we cloned the first cyanobacterial psbO gene, the secondary analysis of the protein indicated a rather expanded protein with a large proportion of ß-sheet structure. Recent hydrodynamic measurements indicated that MSP is elongated, with an axial ratio of ~4:1 (Zubrzycki, et al., 1998). From this perspective, it may not be surprising that modification of function can arise from mutations throughout most of the protein. Our initial site-directed mutations were aimed at charged residues in loops, which we correctly hypothesized were involved with PSII interaction. Mutations within the first N-terminal 30 residues indicated that this region was in proximity, and possibly was involved with binding, to PSII. Much of the work involving modification of MSP amino acids, either by mutation in our lab or by biochemical modification, allowed Bricker and Frankel (1998) to develop a reasonable secondary structure model of MSP (Fig. 17). This model is most valuable for showing which parts of the molecule are likely to be closest to PSII in the membrane and which are interacting with the bulk solvent in the lumen. This model is specifically based on the spinach sequence and is similar to the one we have used for cyanobacteria for a number of years. However, our sequence analysis indicates some specific differences in the cyanobacterial proteins which are different from that of the spinach (Fig. 18).
2. MSP structural analysis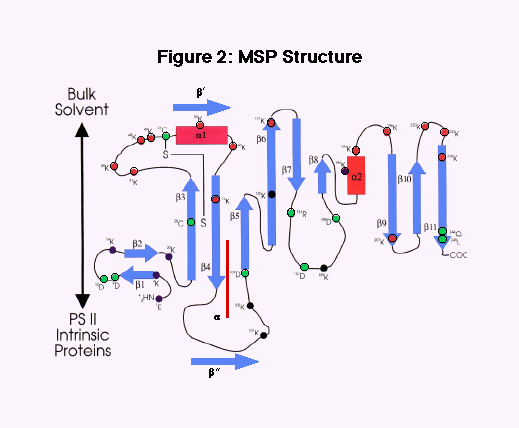{kind=link}
{kind=link}
The MSP sequence of Cyanothece has provided substantial information in analyzing the structure of cyanobacterial MSP. Analysis of the cyanobacterial mutations and the comparison of the sequences allow for the following conclusions, as shown in Fig. 17 and Fig. 18.
| i. | The cyanobacterial and chloroplast proteins are somewhat distinct, particularly in the N-terminal 100 amino acids. Specific lysines are found in the chloroplast, but not in the cyanobacterial protein. |
| ii. | Cyanothece MSP is most closely related to the Synechocystis protein, but there are distinct differences, especially in the regions of amino acid 30-50 from the N-terminus See Fig. 18). |
| iii. | This secondary structural analysis indicates that the cyanobacterial MSP has a ß structure in place of the helical a 1, an a-helical region in place of part of ß 4 and an additional ß in place of part of the large loop between amino acids ~90-110. On the other hand, the C-terminal 150 amino acids give rise to identical structures in both cyanobacteria and chloroplasts. Most importantly, the loop between the ß 3 and a 1 regions are substantially different in cyanobacteria relative to chloroplasts. These differences are highlighted in Fig. 2 by placing additional structural elements near that of the spinach model. |
| iv. | The cyanobacteria lack all four lysines in the 40-50 region and in turn have more negatively-charged amino acids. This is particularly true of Cyanothece which differs most substantially from all other MSPs precisely in this region, and has many more negatively-charged amino acids. We postulate that this region, which is close to the PSII binding region near the N-terminus, is of importance in the overall regulation of O2 evolution. This region may also be directly involved in the conformational changes and that may be involved in the regulation of O2 evolving activity throughout the Cyanothece diurnal cycle. |
To learn more about this work, follow this link.
Back to Sherman Lab Homepage