
A. Research Summary and Background
1. OverviewThe research in this lab is aimed in an understanding of photosynthesis and the structure and biogenesis of the photosynthetic membrane. We utilize prokaryotic cyanobacteria as model organisms because they possess a multitude of important characteristics, including oxygenic photosynthesis similar to plants and the ability to be transformed by DNA.
A major focus at the current time concerns the control of photosynthesis by environmental alterations. These studies began during our research on the unicellular, diazotrophic cyanobacterium, Cyanothece sp. ATCC 51142 which displays a circadian regulation of photosynthesis and N2 fixation. Both photosynthesis and nitrogenase activity showed distinct circadian rhythms with peaks every 24 hours and which were 12 hours out of phase from each other. Our results led us to conclude that metabolism in Cyanothece was controlled through the redox poise of the plastoquinone (PQ) pool. The interrelationship of respiration and photosynthesis changes the redox state of the PQ pool and this has both short-term and long-term effects on cellular processes.
To study this regulation in more detail, we have made use of the transformable, unicellular cyanobacterium, Synechocystis sp. PCC 6803 for which the entire genome has recently been sequenced. This sequence enabled us to identify two gene clusters which resemble genes that have been identified as being involved with redox control in photosynthetic bacteria. We have cloned these genes and made appropriate knockout mutations to determine the effect on photosynthesis and nitrogen metabolism. Our initial studies have indicated that one set of genes affects Photosystem II, whereas the other set of genes is responsible for transcriptional regulation in Photosystem I. We have also isolated a series of other mutations in genes which code for proteins in photosynthesis, in respiration and in nitrogen metabolism, and we can now study the way in which these genes are regulated by the environment. Thus, we can grow the wild-type and mutant cells under iron-deficient and nitrogen-deficient conditions, under high and low light, high and low CO2 concentrations and using a variety of inhibitors which affect redox poise. By comparing the impact of these conditions on the wild-type and mutant strains, we should develop a much better understanding of the genetic regulation of these metabolic processes.
We will also use new technologies to study the global analysis of gene expression in Synechocystis sp. PCC 6803. We are constructing a differential expression library using customized amplification libraries (DECAL). This procedure makes it possible to analyze global transcriptional patterns in prokaryotes because this library removes the abundant rRNA species, among other valuable attributes. With these various tools, we will be able to study the biophysics and biochemistry of photosynthesis along with an in-depth analysis of the regulation of these processes. Such studies will provide new information on the way in which photosynthetic microbes respond to factors such as high or low CO2 concentrations when either iron or nitrogen is limiting and relate to important findings under natural conditions.
2. Recent Research FindingsThe research in this lab has involved the O2-evolving mechanism and the structure and assembly of the photosystems. One recent focus was the study of the lumenal proteins of the O2-evolving complex (especially MSP, the manganese-stabilizing protein), but has progressively moved towards an understanding of the control of photosynthesis. The progression from stage to stage was quite natural and interconnected. Our initial interests were a better understanding of the role played in oxygen evolution by the lumenal proteins of Photosystem II. We have determined that the unicellular cyanobacterium, Synechocystis sp. PCC 6803, could grow photosynthetically in the absence of the psbO gene (encoding MSP). The strain evolved oxygen at about 1/2 to 2/3 the rate of wild-type cells, but did so with very specific alterations, especially in the presence of short, repetitive flashes. At the same time, we were investigating a unicellular, diazotrophic cyanobacterium, Cyanothece sp. ATCC 51142, which had a very interesting type of regulation of photosynthesis and N2 fixation. Both photosynthesis and nitrogenase activity showed distinct circadian rhythms with peaks every 24 hours that were 12 hours out of phase from each other. We determined that oxygen evolution changed quite dramatically throughout the 24-hour period and, at times, our O2-evolving measurements in Cyanothece sp. ATCC 51142 very closely resembled the DpsbO mutation in Synechocystis. This then led us to investigate psbO and O2 evolution in Cyanothece in greater detail. In turn, these studies led us to conclude that Cyanothece demonstrated rather robust state transitions throughout the course of the diurnal cycle and that the control of the state transitions was involved with the regulation of PSII. This led us to determine that the genes encoding the lumenal proteins, such as MSP and the 12 kDa protein (encoded by psbU), were under strong transcriptional regulation. Further studies led us to conclude that all of these properties in Cyanothece were controlled through the redox poise of the plastoquinone (PQ) pool. This hypothesis has been invoked on many previous occasions, but definitive proof was lacking. Thus, we began analyzing the Synechocystis sp. PCC 6803 genome to look for proteins that are homologous to the prrA/B (regA/B) genes known to regulate redox control in photosynthetic bacteria. We have preliminary results that indicate that we have detected such genes and produced appropriate knockout mutations. These results represent one basis for our current work, which analyzes the impact of environmental stresses on photosynthesis and CO2 fixation.
3. PhotosynthesisThe process of photosynthesis encompasses two quite different set of reactions that can be grouped into two broad categories: 1.) photosynthetic electron-transfer reactions (the "light reactions") in which energy derived from sunlight can energize an electron in chlorophyll and enable the electron to move along an electron transport chain until NADP is reduced to NADPH. Since the chlorophyll obtains its electrons from water, the water is split with the concomitant liberation of O2. During the electron transport process, H+ is pumped across the membrane and the resulting proton-motive force drives the synthesis of ATP. Thus, the light reactions result in the production of energy and reducing power, as well as the evolution of O2; 2.) carbon-fixation reactions (the "dark reactions") in which the ATP and NADPH produced by the photosynthetic electron transport reactions help drive the conversion of CO2 to carbohydrate. This lab has been specifically concerned with the light reactions and, in recent years, has concentrated on a better understanding of the O2-evolving mechanism.
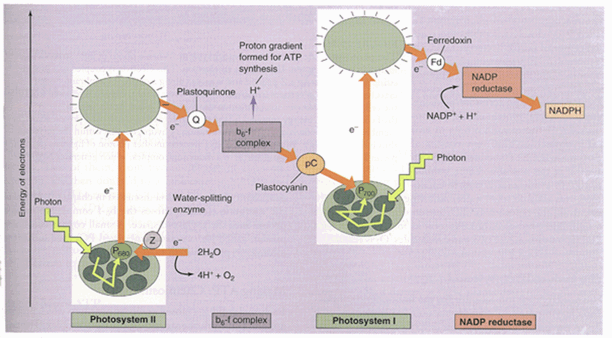
In plants, eukaryotic algae and cyanobacteria, there are two photosystems which are directly excited by light. Each photosystem contains a reaction center in which the energy transduction takes place, along with an antenna which helps funnel light energy toward the reaction center. A scheme showing the redox potential of the various components is referred to as the Z-scheme (Fig. 1). When electrons go all the way from water through PSII, the cytochrome b6/f complex, through PSI and finally to NADP, the process is called non-cyclic electron flow. An alternative pathway of electrons around PSI is termed cyclic electron flow and is involved solely with the production of ATP.
{kind=link}
Altogether, there are four major protein complexes that are embedded in the membrane: Photosystem I and Photosystem II; cytochrome b6/f complex; and ATP synthase. These complexes are highly conserved among all green organisms. However, these organisms differ in the type of antenna systems. Chloroplasts of green plants contain light harvesting complexes which are embedded in the membrane, whereas cyanobacteria have phycobilisomes which are located on the external surface of the membrane (Fig. 2). This lab has been involved with the determination of the proteins associated with each complex and the localization of the complexes within the cyanobacterial thylakoid membranes.
4. Cyanobacteria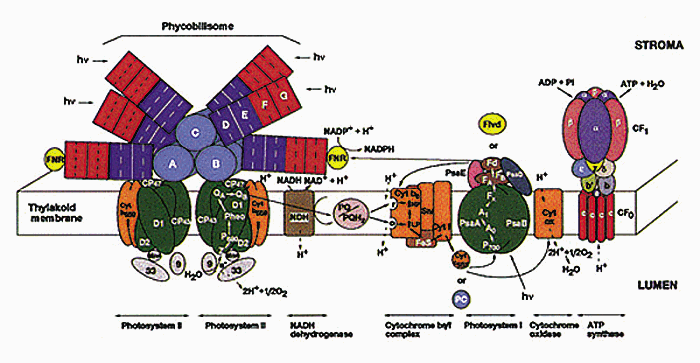{kind=link}
The unicellular cyanobacteria have proven to be excellent model organisms for the study of photosynthesis and, more recently, N2-fixation. Synechococcus sp. PCC7942 and Synechocystis sp. PCC6803 have become among the most valuable of these model systems for the study of the light reactions in photosynthesis because they are capable of genetic exchange via transformation or conjugation. Synechocystis is most valuable for the study of mutations in the photosynthetic apparatus because it is capable of photoheterotrophic growth. Cyanothece is an even more robust organism and can be grown heterotrophically in complete darkness. However, we have yet to demonstrate high frequency gene exchange in this organism.
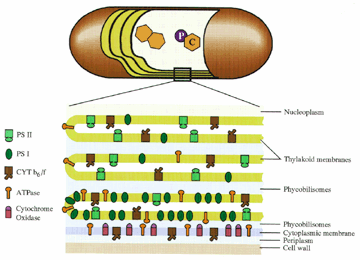
Over the years, we have utilized Synechococcus sp. PCC7942 to the greatest extent and it is the best characterized. It is an autotrophic strain with concentric photosynthetic lamellae (Fig. 3). We have determined the localization of many membrane proteins and have determined that the respiratory apparatus is located in the cytoplasmic membrane. At the same time, the photosynthetic apparatus is not uniformly localized throughout the thylakoids. Rather, PSII is disproportionately high in the inner two thylakoids, whereas PSI is localized to a great extent in the outer thylakoid. Similarly, the ATPase mostly follows the localization of PSI (Fig. 4). We have termed this phenomenon radial asymmetry and find this rigid organization only in Synechococcus. Strains such as Synechocystis sp. PCC6803 (Fig. 5) and Cyanothece (Fig. 6), which have a polar distribution of thylakoids, do not show the same type of asymmetry.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
One very important attribute of cyanobacteria is the plasticity of their metabolic systems. Like other bacteria, they have become adapted for growth under a variety of environmental conditions. We have investigated the photosynthetic apparatus and growth of these cyanobacteria under various environmental conditions, including iron and N2-deficiency, changes in CO2 concentration, as well as various light regimes. This has enabled us to study gene regulation of the various metabolic pathways and we have emphasized this approach in the unicellular, diazotrophic Cyanothece.
5. Photosystem II -- Lumenal side proteinsO2 evolution remains one of the unsolved mechanisms in photosynthesis. The process involves four Mn atoms, which are liganded to the D1 protein of the PSII reaction center (the D1 domain that binds Mn protrudes into the thylakoid lumen). On the lumenal side of the membrane are three extrinsic proteins which are known to bind to the reaction center. Both chloroplasts and cyanobacteria have a protein encoded by the psbO gene that is referred to as the Mn-stabilizing protein (MSP). However, chloroplasts and cyanobacteria differ in the nature of the other two proteins: chlorplasts have a 24 and a 17 kDa protein, whereas cyanobacteria have a cytochrome C550 and a 12 kDa protein.
The function of all of these proteins is still only poorly understood. MSP is needed for optimal rates of H2O-splitting activity, but is not absolutely essential. We have constructed a mutant strain of Synechocystis sp. PCC 6803, in which the entire psbO gene had been deleted. Careful analysis of the deletion mutant demonstrated that it could grow photosynthetically and evolve oxygen at about 60% of the rate of wild-type cells. Thus, MSP is not essential for O2 evolution. However, it is involved in regulation and we have detected its importance in our analysis of Cyanothece sp. ATCC 51142. The role of the other two extrinsic proteins has still not been determined. However, a double mutant that is missing both MSP and cytochrome C550 shows extremely poor O2 evolution. Thus, it would seem that the loss of one protein debilitates PSII O2-evolving capability, but the loss of two of these proteins is more destructive. It is thought that the proteins provide an appropriate environment for the O2-evolving complex and work to elucidate this structure continues.
6. Localization of Membrane ProteinsLocalization of membrane proteins in the cyanobacterium Synechococcus sp. PCC 7942 was determined by transmission electron microscopy utilizing immunocytochemistry with cells prepared by freeze-substitution. This preparation procedure maintained cellular morphology and permitted detection of cellular antigens with high sensitivity and low background. Synechococcus sp. PCC 7942 is a unicellular cyanobacterium with thylakoids organized in concentric layers toward the periphery of the cell. Cytochrome oxidase was localized almost entirely in the cytoplasmic membrane, whereas a carotenoprotein (P35) was shown to be a cell wall component. The major Photosystem II (PSII) proteins (D1, D2, CP43, and CP47) were localized throughout the thylakoids. Proteins of the Cyt b6/f complex were found to have a similar distribution. Thylakoid lumenal proteins, such as the Mn-stabilizing protein, were located primarily in the thylakoid, but a small, reproducible fraction was found in the outer compartment. The Photosystem I (PSI) reaction center proteins and the ATP synthase proteins were found associated mostly with the outermost thylakoid and with the cytoplasmic membrane. These results indicated that the photosynthetic apparatus is not evenly distributed throughout the thylakoids. Rather, there is a radial asymmetry such that much of the PSI and the ATPase synthase is located in the outermost thylakoid. The relationship of this structure to the photosynthetic mechanism is discussed. It is suggested that the photosystems are separated because of kinetic differences between PSII and PSI, as hypothesized by H.-W. Trissl and C. Wilhelm (1993).
A highly schematic model of our findings is represented in the model which is based on the earlier cyanobacterial membrane models from other workers, including Gantt (1980) and Nierzwicki-Bauer et. al (1983). Our results suggest that the cytoplasmic membrane has a complete respiratory chain, including Cyt oxidase, a Cyt b6/f-like complex, and an ATP synthase. The importance of this respiratory pathway as a major metabolic source in Synechococcus strains remains to be determined. The most unique interpretation of our results is that the thylakoid leaflets do not have an identical composition. Instead, the thylakoid closest to the cytoplasmic membrane has a higher percentage of PSI and ATP synthase compared to the inner thylakoids. We believe that the individual membrane sacs have fenestrae and are appressed, thus permitting movement of molecules through the cytoplasm or continuity of the lumenal space for development of pH gradients. The complexes are depicted as randomly distributed in any particular membrane, a feature that obviates the need to develop a mechanism for complex-complex interaction in the plane of the membrane. It is even possible that functional and structural interactions between complexes occur in the thylakoid lumen.
The localization of PSI and ATP synthase, referred to as radial heterogeneity, is similar to the lateral heterogeneity seen in chloroplasts. We can even hypothesize that this organization first developed in cyanobacteria and eventually became the lateral heterogeneity of chloroplasts. Since it is very likely that the thylakoids in cyanobacteria are interconnected, we likely have a more extreme form of lateral heterogeneity. A physical reason for this separation was proposed recently by Trissl and Wilhelm (1993), based on the kinetic differences between PSI and PSII. The authors postulate that PSII is a slow photosystem, whereas PSI is a fast photosystem. Interconnection of the two photosystems could drain off the excitation energy from PSII because PSI has long-wavelength-absorbing pigments and faster trapping kinetics. In cyanobacteria, with a very low PSII:PSI ratio (0.3-0.5), physical separation may be required to prevent this problem.
It should be noted that these results are significant only for cyanobacteria, such as Synechococcus sp. PCC 7942, with concentric thylakoids. We have performed similar experiments on strains, such as Synechocystis sp. PCC 6803 and Cyanothece sp. ATCC 51142, that have thylakoids that protrude radially into the cytoplasm from two or three poles along the cytoplasmic membrane. In these cells, the PSI and ATPase antibodies act quite similarly to the PSII and cytochrome b6/f antibodies and evenly decorate the thylakoids. These cyanobacteria may have a different form of localized compartmentalization, because each of the thylakoids contains a more equal distribution of the photosynthetic complexes. Importantly, the cytochrome oxidase antibodies localize to a much greater extent on the thylakoids in Synechocystis and more evenly over both the thylakoid and cytoplasmic membrane in Cyanothece. These results indicate that the localization of the respiratory chain in cyanobacteria may reflect the nature of the metabolism. Strains, such as Synechocystis and Cyanothece, that are capable of both photoheterotrophic and heterotrophic growth, have cytochrome oxidase in the thylakoid membrane, whereas photoautotrophic organisms, such as Synechococcus, have their cytochrome oxidase located only in the cytoplasmic membrane.
To learn more about this work, follow this link.Back to Sherman Lab Homepage