
F. IMPACT OF ALTERED FE AND NITROGEN CONCENTRATION ON PHOTOSYNTHESIS
The way in which changes in global climate relate to the subsequent response of biospheric photosynthesis is a critical area of study. However, a change in any specific parameter, such as CO2 concentration, is interdependent with other environmental factors, including light, temperature and nutrient supply. This complexity was acknowledged, although poorly understood, until a recent study (Behrenfeld and Kolber, 1999) shed considerable light on the subject. Behrenfeld and Kolber (1999) utilized relatively simple photosynthetic measurements, based primarily on fluorescence, that allowed them to demonstrate the growth of phytoplankton under various nutrient-depleted conditions. In particular, they described some easily observable diel patterns in photosynthetic parameters that were unique to iron-limited phytoplankton, whereas phytoplankton growing in relatively nitrogen-depleted conditions demonstrated somewhat different photosynthetic characteristics. They were then able to duplicate such findings in the laboratory on marine Synechococcus sp. They concluded from their results that primary productivity is iron limited throughout much of the South Pacific gyre. At the same time, the North Atlantic contains sufficient Fe, but is relatively nitrogen deficient and growth and fluorescent properties are different. This study is important for a number of reasons: i.) it highlighted the importance of Fe deficiency on phytoplankton growth; ii.) it showed that similar photosynthetic parameters could be determined in the ocean and in the laboratory; and iii.) it explained these phenomena utilizing the fundamental properties of the photosynthetic mechanism.
Their explanation depends critically on the way in which photosynthetic microorganisms can modify the ratio of Photosystem I (PSI) and Photosystem II (PSII) based on changes in environmental conditions. This is an important element in their explanation of the different fluorescent patterns they obtain for Pacific and Atlantic phytoplankton. In the South Pacific at sunset, Fe limitation results in a decrease of components of the cyt b6/f and PSI complexes, such that the plastoquinone (PQ) pool becomes reduced and induces a transition to what is known as state 2. This is a low PSI/PSII state which prevents complete association of the antenna complexes to PSI. Thus, antenna complexes, which are decoupled from PSII but which are not associated with PSI, can emit absorbed excitation energy which is seen as high levels of background fluorescence. At sunrise, PSI is now more active and can reoxidize the PQ pool, leading to state 1 and a recovery back to the normal fluorescence characteristics. They also propose that Fe concentration in the Atlantic ocean can support relatively high PSI/PSII ratios and that fluorescence changes are more modest, as is found in nitrate-limited Synechococcus. Therefore, they conclude that the differing fluorescence patterns that they observed are due to a redox-controlled mechanism of photosynthesis mediated through the redox poise of the PQ pool. Thus, the growth of prokaryotic, and also likely eukaryotic, photosynthetic microorganisms demonstrates specific photosynthetic characteristics upon growth in different nutrient-deficient conditions. In turn, these characteristics can be described utilizing the unifying features of PQ redox poise and the concept of state transitions.
1. Redox Control of MetabolismIn a very thoughtful review by Geiger and Servaites (1994) on diurnal regulation of photosynthetic carbon metabolism, three levels of regulation were stated: transcription and translation; enzyme activation; and enzyme catalysis (going from the longest to the shortest time frame). Redox control by the PQ pool has the capability of controlling at all three levels. In cyanobacteria, rapid changes might be manifest by state transitions and the movement of phycobilisomes from PSII to PSI (seconds or less). Medium term may involve the build-up or utilization of carbohydrate or nitrogen in storage granules (seconds to minutes). Finally, the longer term control (minutes) may involve the synthesis and degradation of proteins as we have so graphically demonstrated for nitrogenase in Cyanothece (Colón-López et al., 1997). Most of the research performed to date has involved changes in light intensities and a phenomenon known as photoinhibition. However, if this redox control is of general significance, other environmental parameters should also involve redox control. Maxwell et al. (Maxwell et al., 1994, 1995; Savitch et al., 1996) have used temperature, as well as light intensity, to demonstrate the importance of what they call PSII excitation pressure. They conclude that PSII excitation pressure may reflect redox regulation of photosynthetic acclimation to light and temperature in Chlorella. However, their results indicate that the redox state of QA in PSII may only be one of a number of components of a multi-component redox-sensing/signaling pathway. We strongly concur with this conclusion since we feel that respiration, carbon fixation and nitrogen metabolism all play a role in regulating redox poise in cyanobacteria.
Fig. 19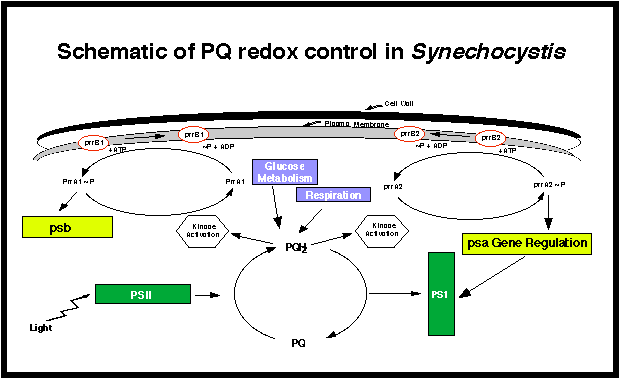{kind=link}
The schematic above briefly summarizes the major metabolic interactions we plan to investigate in the current proposal. The redox state of the PQ pool is central to the story, since it is involved with both photosynthesis and respiration, as well as potential activation of protein kinases. We hope to eventually develop a scheme for cyanobacteria that is as rich in detail as is now available for the adaptation of E. coli to redox environments (Iuchi & Lin, 1993).
2. Redox control in photosynthetic bacteriaThe foundation for this proposal is based on the genetic and molecular research that has been performed in photosynthetic bacteria. The story began with the work of Bauer and colleagues (Mosley et al., 1994) and Eraso and Kaplan (1994, 1995); they identified the prr loci, which were involved in the positive regulation of photosynthesis gene expression in Rhodobacter sphaeroides in response to anaerobiosis. This is a typical two-component regulatory system in bacteria, in that prrB encodes a sensor histidine kinase that is responsive to the removal of oxygen and functions through the response regulator, PrrA (see figure above). PrrA positively regulates the expression of the photosynthesis operons and is thus similar to the RegA gene identified in Rhodobacter capsulatus (Mosley et al., 1994). Thus, in both photosynthetic bacteria, there appears to be a two-component regulatory system that is involved in the transduction of signals that monitor alterations in O2 levels. It is conceivable that redox poise of the PQ pool would be involved in such a process, either as a primary or as a secondary signal. Importantly, Eraso and Kaplan (1996) have determined that mutations in the prrA and prrB genes result in substantially different phenotypes. Mutations in prrA lead to the loss of photosynthesis gene expression, whereas even the absence of prrB does not appear to affect photosynthesis gene expression to a great extent. They postulate that prrB normally phosphorylates prrA, as typical of a two-component regulatory system. However, when prrB is deleted, a heterologous phosphate donor could maintain prrA in the active state. These results suggest that this regulatory operon may be involved in other processes.
The importance of the prrA/B genes was amply demonstrated by Joshi and Tabita (1996). Their work indicated that the prrA/B two-component regulatory system integrates the control of photosynthesis, CO2 assimilation and N2 fixation. In a series of experiments, knockout mutants of prrB were shown to be unable to grow using N2. They demonstrated the interrelationship among photosynthesis, CO2 and N2 fixation. Throughout, they showed the importance of the nitrogenase complex as an alternative redox system for the cell. They conclude that the normal molecular control mechanisms that serve to prevent nif transcription and nitrogenase synthesis in the presence of ammonia are overcome by the organisms¹ need to remove excess reducing equivalents in the absence of a functional carbon fixation cycle. They conclude that the control is somehow mediated by the prrA/B system, and they also highlight the need for the organism to seek various mechanisms to dispense with the large amounts of reducing power generated via photosynthesis and respiration. Once again, we believe that the PQ redox poise is an important cellular signal to integrate the turning on and turning off of these various pathways. It is also important to mention that prrA, the response regulator, shows similarity to patA, a gene described by Haselkorn¹s group to control heterocyst pattern formation in the cyanobacterium Anabaena sp. PCC 7120 (Liang et al., 1992).
3. State TransitionsState transitions were first detected in cyanobacteria by Murata (1969), who noted the effect of light quality on the relative activity of PSI and PSII. Preferential excitation of PSI caused an increase in energy transfer to PSII and a small decrease in energy transfer to PSI, whereas preferential excitation of PSII or incubation in darkness reversed this effect. A model for state transitions in cyanobacteria Fig. 20 has been developed by Rögner and colleagues (Kruip et al. 1994; Rögner et al. 1996; Bald et al. 1996), who have also invoked the oligomeric state of PSI and PSII in the overall mechanism. The crucial point of this model is that there are three different components of state transitions: the attachment of phycobilisomes, the oligomerization of reaction centers, and their organization in the membrane. In this model, state 1 (which favors linear electron flow from O2 production to CO2 fixation) has dimeric PSII organized in rows, a monomeric PSI, and the phycobilisomes are primarily attached to PSII. State 2 (which favors cyclic electron flow) has trimeric PSI complexes, disorganized monomeric PSII, and the phycobilisomes can more readily attach to PSI. State transitions are known to be a major source of non-photochemical quenching of fluorescence in cyanobacteria (Campbell and Öquist 1996). This relationship of oligomerization of the photosystems and state transitions represent an important component of the short-term adaptation in Cyanothece sp. ATCC 51142.
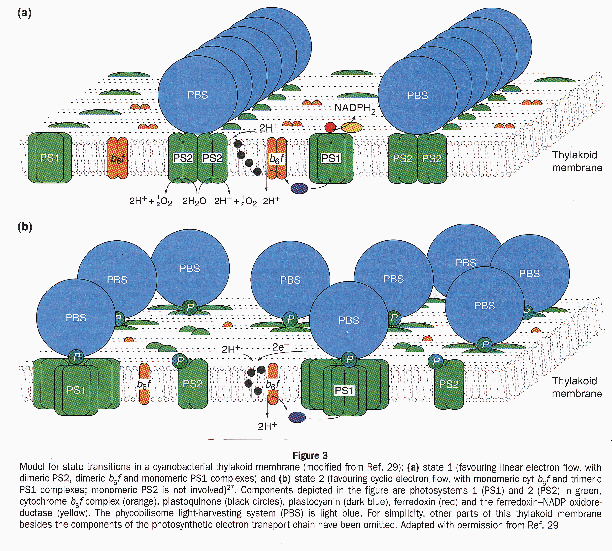{kind=link}
Another model that has been proposed to explain state transitions in cyanobacteria has been termed the "mobile PSI model" (Schluchter et al. 1996). This model is based on the results obtained over a number of years by Bryant and colleagues (Bruce et al. 1989; Shen and Bryant 1995) and is based primarily on results with the psaL deletion mutant which cannot form PSI trimers. Both models postulate the movement of phycobilisomes between PSII and PSI and a specific oligomeric arrangement for PSI.
Two recent papers are of considerable interest and in many ways synthesize years of effort. van Thor et al. (1998) review recent experiments on light harvesting and state transitions in cyanobacteria, including our work in Cyanothece. They come to much the same conclusions, in that phycobilisome complexes are highly mobile on the surface of the photosynthetic membrane. They also propose that the phycobilisomes are in a type of dynamic equilibrium with both photosystems and that regulation of energy transfer is mediated by changes in affinity of the PBS for either photosystem. They don't go into structural details in regard to the oligomeric state of the photosystems, but the conclusions are quite compatible with our own. The second paper (Pfannschmidt et al., 1999) presented the first detailed experiment that utilized both biophysics and molecular biology to show redox control of chloroplast gene expression. They used chloroplast run-on transcription assays to measure mRNA expression, along with the typical fluorescence measurements. They demonstrated convincingly that the redox state of the PQ pool controlled the rate of transcription of genes encoding the PSI and PSII apoproteins. In addition, the stoichiometry between the two photosystems changes in a way that counteracts the inefficiency which is produced when either photosystem limits the rate of the other‹i.e., the state transitions.
4. Nutrient deficiency and the storage of nitrogen and carbonOur research in cyanobacteria has dealt with iron deficiency and nitrogen deficiency and their impact on photosynthesis. Our studies of Fe-deficient growth of cyanobacteria demonstrated substantial changes in the photosynthetic mechanism and that a new Chl-protein was synthesized. Such work represents a component of the fundamental research upon which Behrenfeld and Kolber¹s (1999) conclusions have been built. We have also analyzed the impact of nitrogen deficiency on unicellular, diazotrophic (and non-diazotrophic) cyanobacteria. During this work, we determined that structures are formed in cyanobacteria which are important for the storage of excess nutrient. These structures include cyanophycin granules for the storage of fixed nitrogen and glycogen granules that form under virtually all nutrient-deficient conditions. The formation and degradation of these structures are important for an understanding of the photosynthetic mechanism and carbon flux.
Cyanophycin is a non-ribosomally synthesized, nitrogen-storage polymer in cyanobacteria which is generally composed of the amino acids asp and arg in an equimolar ratio (Simon, 1971, 1987; Allen and Weathers, 1980). In this compound, a poly-aspartate acid backbone is linked to the a amino groups of arg residues through an amide bond formation with the b-carboxyl groups. Cyanophycin has been shown to accumulate in cells which are deficient in specific macro- and micro-nutrients, but which decreases when nitrogen is deficient. Cyanophycin also accumulates when chloromphenicol is added to block protein synthesis; this inhibition causes nitrogenase compounds to accumulate and results in a large increase in cyanophycin and in the corresponding cyanophycin granules (Allen, 1988; Simon, 1971, 1987; Mackerras et al., 1990; Carr, 1988).
In Cyanothece, we see diurnal oscillations in cyanophycin accumulation under N2-fixing conditions. Near the peak of N2 fixation, cyanophycin accumulates in small to medium size granules within the cytoplasm, most of which are localized close to the cytoplasmic membrane. Later in the dark period, the granules grow larger and more numerous as the total amount of cyanophycin in the cell increases. The number and size of cyanophycin granules reaches a peak prior to sunrise and the cyanophycin granules become a large fraction of the total cellular area. The granules then decline throughout the day, reaching a minimum around evening. The importance of cyanophycin in nitrogen metabolism may have been overlooked because of the belief that it was a passive storage compound. However, the results with Cyanothece emphasize the dynamic nature of cyanophycin as a storage reserve in cyanobacteria, as first suggested by Carr (1988). Fortunately, the genes for the cyanophycin synthetase and possibly also for the cyanophycinase have been identified. The synthetase enzyme was isolated from Anabaena variabilis (Ziegler et al., 1998) and the gene determined in Synechocystis sp. PCC 6803. The sequence of this enzyme shows similarities to enzymes of the super family of ligases involved in the biosynthesis of murein and of folyl-poly (g-glutamate). The putative cyanophycinase may have been identified by an interesting mutation in a gene (gcp) that resulted in reduced salt tolerance and enhanced cyanophycin accumulation in Synechocystis sp. PCC 6803 (Zuther et al., 1998). The phenotype included the accumulation of large cyanophycin granules, even under normal growth conditions, but especially after growth in a salt shock of 684 mM NaCl. We suggest that the gcp gene is a candidate for the cyanophycinase and we are making a knockout mutation in this gene, as well as in the synthetase gene.
The impact of different ambient CO2 concentrations on photosynthesis will be another important objective. Recent reviews (Koch, 1996; Drake et al., 1997; Champigny and Gibbs, 1997) have outlined some of the molecular and physiological consequences of increasing CO2 concentrations. Cyanobacteria respond in a related fashion and store carbohydrate in glycogen granules. The inter-relationship of various nutrient alterations is evident ultrastructurally by the accumulation of such granules when cells are grown under Fe or nitrogen deficiency. We will analyze growth, transcription, ultrastructure and biophysical propeties of cells grown under Fe and nitrogen deficiency in the presence of 0.03% and 3.0% CO2 to determine how cyanobacteria respond to changing CO2 concentrations.
To learn more about this work, follow this link.Back to Sherman Lab Homepage